Медицински експерт на статията

Нови публикации
Експериментални модели на остеоартрит
Последно прегледани: 07.07.2025

Цялото съдържание на iLive е медицински прегледано или е проверено, за да се гарантира възможно най-голяма точност.
Имаме строги насоки за снабдяване и само свързваме реномирани медийни сайтове, академични изследователски институции и, когато е възможно, медицински проучвания, които се разглеждат от специалисти. Имайте предвид, че номерата в скоби ([1], [2] и т.н.) са линкове към тези проучвания.
Ако смятате, че някое от съдържанието ни е неточно, остаряло или под съмнение, моля, изберете го и натиснете Ctrl + Enter.
Хрущялът е високоспециализирана тъкан, съдържаща само един вид клетки (хондроцити) и характеризираща се с липсата на кръвоносни и лимфни съдове. Хрущялът се подхранва главно чрез абсорбция от синовиалната течност. Метаболизмът на хондроцитите се регулира от редица разтворими фактори, произвеждани локално от хондроцитите и околните тъкани. Функцията на хондроцитите зависи и от състава на извънклетъчната среда (кислородно напрежение, концентрация на йони, pH и др.), състава на извънклетъчната мембрана (ECM), взаимодействието на клетките и матрицата, както и от физическите сигнали. Основната цел на експерименталното моделиране е да се създадат култури в извънклетъчната среда, без да се променя фенотипът на зрелите клетки. Втората цел е да се създадат култури за изследване на преждевременния, забавен, краткосрочен или продължителен отговор на хондроцитите на химични и/или физически сигнали. In vitro изследванията също предоставят възможност за изследване на поведението на хондроцитите при остеоартроза. Третата цел е разработването на системи за кокултуриране, които позволяват изучаване на взаимодействията на различни тъкани в ставата. Четвъртата задача е подготовката на хрущялни импланти за последваща трансплантация. И накрая, петата задача е изучаването на растежни фактори, цитокини или терапевтични средства, които са способни да стимулират репарацията и/или да инхибират резорбцията на хрущяла.
През последните десетилетия са създадени различни модели на клетъчни култури на ставния хрущял, включително монослойни култури, суспендирани култури, хондронови култури, експлантати, кокултури и безсмъртни клетъчни култури. Всяка култура има своите предимства и недостатъци и всяка е подходяща за изучаване на един специфичен аспект на метаболизма на хондроцитите. По този начин, хрущялните експлантати са отличен модел за изучаване на оборота на матрични елементи, което изисква истински рецептори на клетъчната повърхност и нормални взаимодействия клетка-матрикс и матрица-клетка. В същото време се препоръчва да се изучават матрични отлагания или механизми, регулиращи метаболизма на хондроцитите, върху култура от изолирани клетки. Монослойна култура с ниска плътност е необходима за изучаване на процеса на клетъчна диференциация. Културите, суспендирани в естествена или синтетична матрица, са модел за анализ на адаптивния отговор на хондроцитите към механично натоварване.
Хондроцитни култури
При избора на хрущялна тъкан за in vitro изследвания трябва да се вземат предвид няколко важни момента. Съставът на матрицата и метаболитната активност на хондроцитите варират между ставите, като последното зависи и от дълбочината на разположение на хондроцитите в тъканта. Тези данни са получени в няколко експеримента, в които са изследвани изолирани субпопулации от хондроцити от хрущялни зони с различна дълбочина. Установени са редица морфологични и биохимични разлики между култивирани хондроцити, разположени в повърхностните и дълбоките слоеве на ставния хрущял. Повърхностните клетки синтезират рядка, бедна на протеогликани фибриларна матрица, докато по-дълбоките клетки произвеждат матрица, богата на фибрили и протеогликани. Освен това, повърхностните клетки произвеждат относително повече малки неагрегирани протеогликани и хиалуронова киселина и относително по-малко агрекан и кератан сулфат, отколкото по-дълбоките хондроцити. Друга важна отличителна черта на метаболизма на хондроцитите, изолирани от хрущялни зони с различна дълбочина, е отговорът на екзогенен стимул. Според M. Aydelotte et al., говеждите хондроцити от повърхностната зона на хрущяла са по-чувствителни към IL-1, отколкото клетките от дълбоката зона.
Клетъчното поведение зависи и от местоположението на тъканта. Хондроцитите от хрущял на ребрата и ухото от едно и също животно реагират различно на растежни фактори като фибробластен растежен фактор (FGF) и TGF-бета. FGF увеличава включването на тимидин, пролин и левцин в култивираните хондроцити на ребрата, но не и в ухото. TGF-бета увеличава включването на тимидин в хондроцитите на хрущяла на ребрата и ухото, но не оказва влияние върху включването на тимидин и пролин в ухото. Хрущялните клетки от области с висок стрес се различават от тези от области с нисък стрес върху хрущяла. По този начин, хондроцитите на зрял хрущял на колянната става на овце от централната област на ставната повърхност на пищяла, която не е покрита от менискуса, който носи най-голямо натоварване in vivo, синтезират по-малко агрекан, но повече декорин, отколкото клетките от зоните, покрити от менискуса. Авторите също така подчертават важността на използването на хрущял от идентични ставни зони при изучаване на синтетичната функция на ставите.
Метаболизмът на хондроцитите и техният отговор към регулаторни фактори също зависят значително от възрастта на донора, неговото скелетно развитие и състоянието на ставите, от които са взети клетките. При човешките хондроцити се наблюдава значително намаляване на пролиферативния отговор с възрастта. Най-голямо намаление се наблюдава при донори на възраст 40-50 години и над 60 години. Освен това, тежестта на пролиферативния отговор към растежни фактори (напр. FGF и TGF-бета) намалява с напредване на възрастта. В допълнение към количествените промени в пролиферацията на хондроцитите, има и качествени промени. Клетките от млади донори (10-20 години) реагират по-добре на тромбоцитен растежен фактор (PDGF), отколкото на TGF-бета, докато обратното се наблюдава при клетки от възрастни донори. Използват се няколко механизма, за да се обяснят възрастово зависимите промени в синтетичната функция на хондроцитите и техния отговор към растежни фактори. Те включват намаляване на броя и афинитета на повърхностните клетъчни рецептори, промени в синтеза и биоактивността на растежните фактори и цитокините и модификация на пострецепторните сигнали.
Патологичното състояние на ставите също променя морфологията и метаболитната активност на хондроцитите. Така, J. Kouri et al. (1996) идентифицират три субпопулации от хондроцити в хрущяла при остеоартроза. Хондроцитите от повърхностната и горната част на средната част на хрущяла образуват клъстери и синтезират по-голямо количество протеогликани и колаген. TGF-бета и инсулиноподобният растежен фактор (IGF) са способни да стимулират синтеза на протеогликани от хондроцитите и частично да неутрализират ефектите на IL-1 и TNF-a. Експлантати на хрущял, засегнати от остеоартроза, и хондроцити, изолирани от хрущяла на пациент с остеоартроза, са по-чувствителни към стимулация на TGF-бета, отколкото хондроцитите на здрав хрущял. Тези разлики най-вероятно са свързани с фенотипни промени в хондроцитите в горните слоеве на ставния хрущял.
Изолирането на отделни хондроцити се постига чрез последователно третиране на извънклетъчната мембрана (ECM) с протеолитични ензими. След освобождаването им от ECM, изолираните клетки са идеални за изучаване на de novo синтеза на матрични компоненти. Някои автори използват само клостридиална колагеназа, докато други предварително инкубират хрущяла с трипсин, проназа, ДНКаза и/или хиалуронидаза. Броят на изолираните клетки зависи от използваните ензими. Така, когато се третират само с колагеназа, от 1 g тъкан могат да се получат 1,4-10⁶ хондроцити, докато при използване на проназа, хиалуронидаза и колагеназа -4,3-10⁶. Когато се третират с колагеназа, агрекан, протеини, IL-6 и IL-8 остават в клетъчната култура в значително по-големи количества, отколкото при последователно третиране с различни ензими. Има няколко обяснения за тези разлики между двете клетъчни култури:
- Клетъчните рецептори са увредени или инхибирани от ензими, TGF-бета инхибира синтеза на ДНК и протеогликани в прясно изолирани хондроцити (ден 1), докато синтезът на ДНК и протеогликани в хондроцити, култивирани в монослой (7 дни), се стимулират от TGF-бета. Необходим е обаче адекватен период за повторна експресия на тези мембранни компоненти преди началото на експеримента.
- Екзогенните протеази могат да нарушат интегрин-медиираното взаимодействие между клетката и матрикса. Семейството интегрини насърчава прикрепването на хондроцитите към молекулите на извънклетъчната матриксна мембрана (Shakibaei M. et al., 1997). Това нарушаване може да повлияе на експресията на матрични гени.
- Остатъците от матрични компоненти могат да регулират синтетичната функция на хондроцитите. Интегрините са способни да разпознават продуктите от разграждането на извънклетъчната мембрана (ECM), като по този начин играят важна роля в тъканната репарация след действието на протеолитични ензими. Т. Ларсон и др. (1989) съобщават, че добавянето на непокътнати или фрагментирани протеогликани към клетъчната култура стимулира синтеза на протеини и протеогликани. Високото ниво на хиалуронова киселина обаче води до значително намаляване на включването на сулфати в синтеза на протеогликани от хондроцитите на пилешкия ембрион, зрелите хондроцити на прасешките и плъши хондросаркомни клетки. Освен това, хиалуроновата киселина е инхибитор на освобождаването на протеогликани от клетките дори в присъствието на IL-1b, TNF-a, FGF, което показва противодействие на първата биологична активност на растежните фактори и цитокините. Точният механизъм, който стои в основата на действието на хиалуроновата киселина, остава неясен; Известно е, че хондроцитите съдържат рецептор за хиалуронова киселина, свързан с актинови филаменти на цитозола. Свързването на хиалуроновата киселина с нейния рецептор стимулира фосфорилирането на протеини. Следователно, тези данни демонстрират модулация на метаболитната функция на хондроцитите от фрагментирани или нативни молекули на матрични протеини чрез активиране на рецепторите на клетъчната мембрана.
- Бързото стимулиране на синтеза на матрични протеини от хондроцитите чрез ензими може да е следствие от промени във формата на хондроцитите и/или реорганизация на цитоскелета.
- Някои цитокини (напр. IL-8) и растежни фактори (напр. IGF-1, TGF-β) са секвестирани в извънклетъчната мембрана (ECM). Най-известният пример е свързването на TGF-β с декорин, което води до намалена способност на първия да индуцира клетъчен растеж в яйчникови клетки на китайски хамстер. Констатацията, че съдържанието на декорин в хрущяла се увеличава с възрастта, предполага намаляване на бионаличността на TGF-β с напредване на възрастта. Растежните фактори и цитокините могат да се освобождават от матричните остатъци по време на култивирането и впоследствие да модулират функцията на хондроцитите.
Монослойна култура от хондроцити
Диференцираният фенотип на хондроцитите се характеризира предимно със синтеза на колаген тип II и тъканно-специфични протеогликани, както и с ниско ниво на митотична активност. Има данни, че при продължително култивиране на клетки в монослой, както и след няколко повторни клетъчни пасажа, хондроцитите губят сферичните си очертания и придобиват удължена, фибробластоподобна форма. При такава фибробластна метаплазия се променя и синтетичната функция на клетките, характеризираща се с прогресивно намаляване на синтеза на колагени типове II, IX и XI и увеличаване на синтеза на колагени типове I, III и Y. Малки неагрегирани протеогликани се синтезират поради функционален агрекан. Синтезът на катепсин B и L е изключително нисък в диференцираните клетки, но се увеличава в процеса на загуба на диференциация. Колагеназа-1 се експресира в диференцираните хондроцити; при продължително култивиране експресията ѝ намалява, докато производството на тъканни инхибитори на металопротеиназите (TIMPs) се увеличава.
Диференцираните хондроцити реекспресират колаген от диференцирания фенотип, когато се прехвърлят от монослойна в суспендирана култура. Процесът на диференциация вероятно е свързан с формата на клетките. Това свойство се използва редовно от изследователи, изучаващи дефектни присадки с автоложни хондроцити. Малък брой клетки, получени от биопсичен материал, могат да бъдат разширени в монослойна култура и след това отново въведени в триизмерна матрица преди трансплантация. Реекспресията на специфичен фенотип от дедиференцирани хондроцити, прехвърлени в агарозна култура, може да бъде стимулирана от TGF-β, осеин-хидроксиапатитен комплекс и аскорбинова киселина.
В отговор на растежни фактори и цитокини, хондроцитите се модифицират по време на процеса на диференциация. Клетъчният отговор към цитокините и растежните фактори се различава между недиференцираните и диференцираните хондроцити. IL-1 стимулира пролиферацията на фибробластите, докато растежът на недиференцираните хондроцити се инхибира от IL-1. Синтезът на ДНК се стимулира от IGF-1 в удължени, но не сплескани хондроцити. В диференцираните хондроцити стимулиращите ефекти на IL-1β и TNF-α върху производството на проколагеназа са по-изразени, отколкото в недиференцираните хондроцити.
Култивиране на хондроцити
Култивирането на хондроцити в суспензия в течна среда или в естествена или синтетична триизмерна матрица стабилизира хондроцитния фенотип. Клетките запазват сферичната си форма и синтезират тъканно-специфични протеини. Суспендираната култура от хондроцити обикновено се препоръчва за изследване на образуването на нова перицелуларна матрица. Културите от хондроцити в синтетични или естествени абсорбиращи полимери се използват за имплантиране на клетки в хрущялни дефекти, за да се стимулира регенерацията на ставната хрущялна тъкан. Синтетичната или естествената среда за имплантирани клетки трябва да отговаря на редица изисквания:
- имплантите трябва да имат пореста структура за клетъчна адхезия и растеж,
- нито самият полимер, нито продуктите от неговото разграждане трябва да причиняват възпаление или токсични реакции при имплантиране in vivo,
- Носителят на присадката трябва да има способността да се свързва със съседен хрущял или субхондрална кост,
- естествената или синтетичната матрица трябва да има способността да абсорбира, нейното разграждане трябва да бъде балансирано чрез регенерация на тъканите,
- За да се улесни възстановяването на хрущяла, химическата структура и архитектурата на порите на матрицата трябва да улесняват поддържането на клетъчния фенотип и синтеза на тъканно-специфични протеини от хондроцитите, разположени в нея,
- По време на in vivo имплантация е необходимо да се изследват механичните свойства на синтетичната или естествената матрица.
 [ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
[ 12 ], [ 13 ], [ 14 ], [ 15 ], [ 16 ]
Суспензия на хондроцити в течна фаза
Прикрепването на клетки към пластмасови съдове, в които се култивират хондроцити, може да се предотврати чрез покриване на стените им с разтвор на метилцелулоза, агароза, хидрогел (поли-2-хидроксиетилметакрилат) или смес от колаген и агароза. При тези условия хондроцитите образуват клъстери и синтезират главно агрекан и тъканно-специфични колагени (II, IX, XI типове). Обикновено се откриват два вида клетки. Клетките, разположени в центъра, запазват сферична форма и са обградени от добре развит извънклетъчни матрикс (ECM), което се потвърждава от хистохимични и ултраструктурни изследвания. В периферията хондроцитите имат дисковидни очертания и са обградени от рядък ECM; малко се знае за функционалните характеристики на такива клетки.
Възможно е да се култивират хондроцити върху микроносители, поддържани в суспензия; като микроносители се използват декстранови перли (цитодекс), покрити с колаген декстранови перли (цитодекс III) и непорести микросфери от колаген тип I (целаген). При тези условия на култивиране хондроцитите се прикрепят към повърхността на микроносителя, запазват сферичната си форма и произвеждат матричноподобен материал. Освен това, използването на целлаген насърчава пролиферацията на хондроцитите и повторното експресиране на нормалния фенотип. Следователно, култивирането на хондроцити върху целлагенови микросфери може да се използва за възстановяване на клетъчния фенотип преди трансплантация.
Друг метод за култивиране на хондроцитна суспензия в течна среда е култивирането им под формата на плътни топчета, състоящи се от клетки (0,5-1 * 10 b ), получени чрез центрофугиране. Такива хондроцити са способни да произвеждат матрица, съдържаща голямо количество протеогликани, колаген тип II, но не и колаген тип I, което се потвърждава чрез хистологични, имунохистохимични и количествени методи.
Суспензия на хондроцити в естествен ECM
Хондроцитите могат да се култивират в суспензия в триизмерна матрица (мек агар, агароза, колагенов гел или гъба, хиалуронова киселина, фибриново лепило, алгинатни перли).
Хондроцитите, култивирани в агароза, запазват нормалния си фенотип и синтезират колаген тип II и тъканно-специфични агреканови агрегати. При култивиране в агароза, протеогликаните, синтезирани от клетката, се освобождават в средата в продължение на 50 дни. За сравнение, при монослойна култура клетъчната фаза е препълнена с гликозаминогликани още през първите 5-6 дни от култивирането; при култивиране в средата, след повишен синтез и освобождаване на гликозаминогликани през първите 8-10 дни, настъпва тяхното зависимо от времето намаляване. Въпреки това, поведението на хондроцитите при култивиране в агароза се различава от това in vivo. В агароза голям брой синтезирани агреканови агрегати съдържат по-малки молекули и по-малко молекули, отколкото in vivo. TGF-β стимулира синтеза на протеогликани в експланта, но намалява синтеза на агрекани в агароза.
Алгинатът е линеен полизахарид, получен от кафяви водорасли. В присъствието на двувалентни катиони, като например Ca2 + йони, този полимер се превръща в гел. Всеки хондроцит, хванат в алгинат, е заобиколен от матрица от отрицателно заредени полизахариди, чиито пори са сравними с тези в хиалиновия хрущял. Матрицата, образувана от хондроцитите в алгинатните перли, се състои от две отделения - тънък слой от клетъчно-асоциирана матрица, съответстващ на перицелуларната и териториална матрица на ставния хрущял, и по-отдалечена матрица, еквивалентна на интертериториалната в естествената тъкан. На 30-ия ден от култивирането относителният и абсолютният обем, зает от клетките и всяко от двете отделения в алгинатна перла, са почти напълно идентични с тези в естествения хрущял. В продължение на почти 30 дни хондроцитите запазват сферичната си форма и произвеждат агрекан, чиито хидродинамични свойства са подобни на тези на агрекановите молекули в матрицата на ставния хрущял, както и колагенови молекули от типове II, IX и XI. В същото време, подобно на други суспензионни култури, на повърхността на алгинатни перли присъстват сплескани клетки, които произвеждат малко количество колагенови молекули от тип I, които се освобождават директно в средата и не се включват в ECM. В алгинатните перли се наблюдава умерена пролиферация на хондроцити. След 8 месеца култивиране в алгинатен гел, зрелите хондроцити не губят метаболитна активност и продължават да синтезират тъканно-специфичен колаген тип II и агрекан.
Х. Танака и др. (1984) изследват дифузионните свойства на различни естествени молекули в алгинат и установяват, че молекули по-големи от 70 kDa не дифундират през алгината. Следователно, клетъчната култура в алгинат е подходяща за изучаване на регулацията на матричната биосинтеза и организацията на извънклетъчната матриксна маса (ECM). Наличието на клетки, култивирани в алгинат, позволява да се изследва действието на пептидните регулаторни фактори и фармакологичните агенти на транскрипционно, посттранскрипционно и транслационно ниво.
Хондроцитите също се култивират в матрица от колагенови влакна от типове I и II. S. Nehrer et al. (1997) сравняват функционирането на кучешки хондроцити в порести колаген-протеогликанови полимерни матрици, съдържащи колагени от различни типове. Те откриват важни разлики в морфологията на биосинтетичната функция на хондроцитите, култивирани в колагенови матрици, съдържащи колаген типове I и II. Клетките в матрица от колаген тип II запазват сферичната си форма, докато в колаген тип I имат фибробластоподобна морфология. Освен това, в матрица от колаген тип II, хондроцитите произвеждат по-голямо количество гликозаминогликани. J. van Susante et al. (1995) сравняват свойствата на хондроцитите, култивирани в алгинатен и колагенов (тип I) гел. Авторите откриват значително увеличение на броя на клетките в колагеновия гел, но от 6-ия ден на култивиране клетките губят характерния си фенотип, превръщайки се във фибробластоподобни клетки. В алгинатния гел се наблюдава намаляване на броя на клетките, но хондроцитите запазват нормалния си фенотип. В колагеновия гел броят на протеогликаните на клетка е значително по-висок, отколкото в алгината, но в гела се наблюдава намаляване на синтеза на матрични елементи, започвайки от 6-ия ден на култивиране, докато в алгината синтезът продължава да се увеличава.
Твърдата триизмерна фибринова матрица е естествено вещество, което поддържа хондроцитите, суспендирани в нея, в диференциран фенотип. Триизмерната фибринова матрица може да се използва и като носител при трансплантация на хондроцити. Предимствата на фибрина са липсата на цитотоксичност, способността за запълване на пространството и адхезивният капацитет. Хистологични и биохимични изследвания, авторадиография и електронна микроскопия показват, че хондроцитите във фибринов гел запазват своята морфология, пролиферират и произвеждат матрица дори след 2 седмици култивиране. Въпреки това, G. Homminga et al. (1993) съобщават, че разпадането на фибрина започва след 3 дни култивиране и дедиференциацията на хондроцитите напредва.
Суспензия на хондроцити в изкуствен (синтетичен) ECM
Хрущялни импланти за реконструктивна или ортопедична хирургия могат да бъдат получени чрез отглеждане на изолирани хондроцити in vitro в синтетична биосъвместима матрица.
Хондроцитите, култивирани в полигликолова киселина, пролиферират и поддържат нормална морфология и фенотип в продължение на 8 седмици. Комплексът хондроцити-полигликолова киселина се състои от клетки, гликозаминогликани, колагени и има външна колагенова капсула. Такива импланти обаче съдържат два вида колагенови молекули - I и II. Имплантите от хондроцити, дедиференцирани чрез серия от пасажи, имат по-голямо количество гликозаминогликани и колагени, отколкото имплантите от предимно недиференцирани хондроцити.
Л. Фрийд и др. (1993b) сравняват поведението на човешки и говежди хондроцитни култури във фиброзна полигликолова киселина (FPGA) и пореста полимлечна киселина (PPLA). След 6-8 седмици култивиране на говежди хондроцити в FPGA или PPLA, авторите наблюдават клетъчна пролиферация и регенерация на хрущялната матрица. В FPGA хондроцитите имат сферична форма и са разположени в лакуни, заобиколени от хрущялна матрица. След 8 седмици култивиране in vitro, регенерираната тъкан съдържа до 50% сухо вещество (4% клетъчна маса, 15% гликозаминогликани и 31% колагени). В PPLA клетките имат вретеновидна форма и малко количество гликозаминогликани и колаген. В FPGA клетъчният растеж е 2 пъти по-интензивен, отколкото в PPLA. In vivo, хондроцитите, отглеждани в VPGK и PPLC, произвеждат тъкан, хистологично подобна на хрущяла, в рамките на 1-6 месеца. Имплантите съдържат гликозаминогликани, колагени тип I и II.
Хондроцитите от говежди фетални клетки бяха култивирани в порест хидрофобен и хидрофилен полиетилен с висока плътност. След 7 дни инкубация и в двата субстрата, клетките запазиха сферична форма и съдържаха предимно колаген тип II. След 21 дни култивиране беше установено, че хидрофилната матрица съдържа повече колаген тип II, отколкото хидрофобната матрица.
Хрущялна тъкан може да се получи и чрез култивиране в монослой върху филтри Millicell-CM. Предварителното покритие на филтрите с колаген е необходимо за прикрепването на хондроитините. Хистологичното изследване на културата показва натрупването на хондроцити в извънклетъчния хрущял (ECM), съдържащи протеогликани и колаген тип II. Колаген тип I не е открит в такава култура. Хондроцитите в получената хрущялна тъкан са със сферична форма, но на повърхността на тъканта са донякъде сплескани. Дебелината на новообразуваната тъкан се увеличава с времето и зависи от началната плътност на клетъчния монослой. При оптимални условия на култивиране дебелината на хрущялната тъкан достига 110 μm, организацията на нейните клетки и колаген в повърхностни и дълбоки слоеве е подобна на тази на ставния хрущял. ECM съдържа приблизително 3 пъти повече колаген и протеогликани. След 2 седмици култивиране се наблюдава натрупване на матрикс, което позволява тъканта да бъде извлечена от филтъра и използвана за трансплантация.
Симс и др. (1996) са изследвали култивирането на хондроцити в полиетиленоксиден гел, капсулирана полимерна матрица, която позволява трансфера на голям брой клетки чрез инжектиране. Шест седмици след инжектирането в подкожната тъкан на атимични мишки се е образувал нов хрущял, който морфологично се е характеризирал с бяла опалесценция, подобна на хиалинен хрущял. Хистологичните и биохимичните данни показват наличието на активно пролифериращи хондроцити, произвеждащи извънклеточен матрикс (ECM).
Обяснение
Експлантацията на хрущялна тъкан се използва за изучаване на процесите на ана- и катаболизъм в нея, поддържане на хомеостаза, резорбция и възстановяване. Хондроцитите в хрущялните експлантати поддържат нормален фенотип и състав на извънклетъчния матрикс (ECM), подобни на тези в ставния хрущял in vivo. След 5 дни култивиране в присъствието на серум се постига постоянно ниво на синтез и естествени процеси на разграждане. Резорбцията на тъканите може да се ускори в основната култура и в култура с добавяне на серум, като се използват редица агенти, като IL-IB, TNF-a, бактериални липополизахариди, производни на ретиноева киселина или активни кислородни радикали. За изследване на репарацията на хрущяла, неговото увреждане се индуцира от разтворими възпалителни медиатори (H2O2, IL- 1, TNF-a) или физическо разкъсване на матрицата.
Методът на органотипната култура е модел за in vitro изследвания на ефектите на изолирани външни фактори върху хондроцитите и околната матрица. In vivo хондроцитите са разположени рядко в извънклетъчната мембрана (ECM) и не контактуват помежду си. Културата от експлантиран ставен хрущял запазва тази структурна организация, както и специфичните взаимодействия между хондроцитите и околната извънклетъчна среда. Този модел се използва и за изследване на ефектите на механичното натоварване, фармакологичните агенти, растежните фактори, цитокините и хормоните върху метаболизма на хрущяла.
Друго предимство на експлантацията на хрущялна тъкан е липсата на увреждане на хондроцитите под действието на протеолитични ензими или механични фактори, което е неизбежно при изолиране на клетки. Рецепторите и други мембранни протеини и гликопротеини са защитени от увреждащи фактори.
[ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ]
Хондронна култура
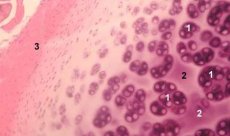
Хондронът е структурна, функционална и метаболитна единица на ставния хрущял, състояща се от хондроцит, неговата перицелуларна матрица и компактна нишковидна капсула и отговорна за матриксната хомеостаза. Хондроните се извличат механично от хрущяла и се събират чрез няколко последователни нискоскоростни хомогенизации. Хондроните, изолирани от зони с различна дълбочина на хрущяла, могат да бъдат разделени на четири категории: единичен хондрон, сдвоени хондрони, множество (три или повече) линейно разположени хондрони (хондронни колони) и хондронови клъстери.
Единични хондрони обикновено се намират в средните слоеве на непокътнатия хрущял, сдвоени хондрони се намират на границата между средния и дълбокия слой, линейно разположените множество хондрони са типични за дълбоките слоеве на непокътнатия хрущял. Накрая, хондроновите клъстери се състоят от произволно организирани групи от единични и сдвоени хондрони, които запазват агрегирано състояние след хомогенизиране. Хондроновите клъстери са големи фрагменти от хрущял, обикновено съдържащи няколко хондрона и радиално разположени колагенови фибрили, т.е. типична организация, характерна за дълбоките слоеве на матрикса. Хондроните са имобилизирани в прозрачна агароза, което позволява изследване на тяхната структура, молекулен състав и метаболитна активност. Хондрон-агарозната система се счита за микромодел на хрущяла, който се различава от традиционната хондроцитно-агарозна система по това, че естествената микросреда е запазена и няма нужда от нейното синтезиране и сглобяване. Хондронната култура е модел за изучаване на взаимодействията на клетките и матрикса в ставния хрущял при нормални и патологични условия.
[ 22 ], [ 23 ], [ 24 ], [ 25 ], [ 26 ], [ 27 ]
Култура на безсмъртни хондроцити
Рекомбинантна ДНК или вируси, съдържащи онкогени, способни да направят клетката „безсмъртна“, се използват за създаване на постоянни клетъчни линии. Безсмъртните хондроцити имат способността да се размножават безкрайно, като същевременно поддържат стабилен фенотип. F. Mallein-Gerin et al. (1995) показват, че онкогенът SV40T индуцира пролиферацията на миши хондроцити, които продължават стабилно да експресират колаген типове II, IX и XI, както и ставен агрекан и свързващ протеин. Такава клетъчна линия обаче придобива способността да синтезира колаген тип I, когато се култивира в монослойна култура или в агарозен гел.
У. Хортън и др. (1988) описват линия от безсмъртни клетки с ниско ниво на експресия на мРНК на колаген тип II. Тези клетки са получени чрез трансформацията им с миши ретровирус, съдържащ I-myc- и y-ra-онкогени. Този тип клетки представляват уникален модел за изучаване на взаимодействията на ставния матрикс при липса на колаген тип II, както и на регулирането на синтеза на колаген тип II.
Култивирането на хондроцити с мутирали или делетирани гени е удобен модел за изучаване на тяхната физиологична функция. Този модел е особено подходящ за изучаване на ролята на специфични молекули в организацията на хрущялната матрица или за изследване на ефектите от различни регулаторни фактори върху метаболизма на хрущяла. Хондроцитите с делетиран ген за колаген тип IX синтезират колагенови фибрили, които са по-широки от нормалното, което показва, че колагенът тип IX регулира диаметъра на фибрилите. Както е отбелязано в Глава 1, наскоро беше открита мутация в гена COLAI, кодиращ колаген тип II, в семейства с първичен генерализиран остеоартрит. За да изследват ефекта на мутантния колаген тип II върху ставната матрица, R. Dharmrvaram et al. (1997) трансфектират (инфектират с чужда нуклеинова киселина) дефектен COL 2 AI (аргининът в позиция 519 е заменен с цистеин) в човешки фетални хондроцити in vitro.
Система за кокултура. В ставата хрущялът взаимодейства с други видове клетки, съдържащи се в синовиалната мембрана, синовиалната течност, връзките и субхондралната кост. Метаболизмът на хондроцитите може да бъде повлиян от различни разтворими фактори, синтезирани от изброените клетки. По този начин, при артрит, ставният хрущял се разрушава от протеолитични ензими и свободни радикали, произвеждани от синовиалните клетки. Поради това са разработени модели за изучаване на сложни взаимодействия между хрущяла и околните тъкани, които се наричат кокултури.
S. Lacombe-Gleise et al. (1995) култивират заешки хондроцити и остеобласти в система за кокултуриране (COSTAR), в която клетките са разделени от микропореста мембрана (0,4 μm), позволяваща обмен между двата типа клетки без директен контакт. Това проучване демонстрира способността на остеобластите да стимулират растежа на хондроцитите чрез разтворими медиатори.
А. М. Малфе и съавтори (1994) изследват връзката между моноцитите в периферната кръв и хондроцитите. Този модел е удобен за изучаване на цитокин-медиирани процеси при възпалителни артропатии (ревматоиден артрит, серонегативни спондилоартрити и др.). Авторите на модела разделят клетките с протеин-свързваща мембрана с пори с диаметър 0,4 μm. Изследването показва, че моноцитите, стимулирани с липополизахарид, произвеждат IL-1 и TNF-α, които инхибират синтеза на агрекан от хондроцитите и допринасят за разграждането на вече синтезираните агрегати на агрекан.
К. Тада и др. (1994) създават модел на кокултура, в който ендотелни клетки в колагенов (тип I) гел са поставени във вътрешна камера, отделена от външната камера с хондроцити, поставени в нея чрез филтър с размер на порите 0,4 μm. В състояние на пълна изолация от външната камера, човешките ендотелни клетки образуват тръбички в колагенов гел в присъствието на EGF или TGF-α. Когато и двата типа клетки се култивират едновременно, образуването на тръбички, зависими от TGF-α, от ендотелните клетки е инхибирано. Инхибирането на този процес от хондроцитите е частично елиминирано от анти-TGF-β антитела. Може да се предположи, че TGF-β, произведен от хондроцитите, инхибира васкуларизацията на самия хрущял.
С. Грут и др. (1994) едновременно култивират хондроцити от хипертрофичните и пролиферативните зони на костта на 16-дневен миши фетус с парченца мозъчна тъкан. След 4 дни култивиране се наблюдава трансдиференциация на хондроцитите в остеобласти и начало на образуване на остеоиди. След 11 дни култивиране част от хрущяла е заменена от костна тъкан, а костната матрица е частично калцифицирана. Някои невропептиди и невротрансмитери, произвеждани от мозъчната тъкан, влияят върху метаболизма на остеобластите или имат рецептори за тях. Сред тях са норепинефрин, вазоактивен чревен пептид, пептид, свързан с гена на калцитонина, субстанция Р и соматостатин. Парчета мозъчна тъкан, култивирани заедно с хондроцити, могат да произведат някои от изброените фактори, способни да индуцират процеса на трансдиференциация на хондроцитите в остеобласти.
[ 28 ], [ 29 ], [ 30 ], [ 31 ], [ 32 ], [ 33 ]
Влиянието на външни фактори върху хондроцитна култура
Влиянието на кислородното налягане върху метаболизма на хондроцитите
В повечето случаи хондроцитните култури се развиват при условия на атмосферно кислородно напрежение. Добре известно е обаче, че in vivo хондроцитите съществуват при хипоксични условия и кислородното напрежение варира при различни патологични състояния. По време на процеса на зреене се наблюдават значителни промени в кръвоснабдяването на епифизите. Тъй като васкуларизацията варира в различните зони на растежната плоча, кислородното напрежение в тях също варира. C. Brighton и R. Heppenstall (1971) демонстрират, че в тибиалната плоча на зайци кислородното напрежение в хипертрофичната зона е по-ниско, отколкото в околния хрущял. Измерванията на някои метаболитни параметри показват, че хондроцитите са способни бързо да реагират на локални промени в концентрацията на кислород. На първо място, при ниско кислородно напрежение, консумацията му от хондроцитите намалява. С намаляване на кислородното напрежение от 21 до 0,04% се увеличава усвояването на глюкоза, увеличава се активността на гликолитичните ензими и синтезът на млечна киселина. Дори при ниско кислородно напрежение абсолютното количество АТФ, АДФ и АМФ остава стабилно. Тези данни показват, че метаболизмът на хондроцитите е насочен към максимално запазване на енергията. Въпреки това, синтетичната активност и следователно процесите на репарация се променят при хипоксични условия.
Високото кислородно налягане също влияе върху метаболизма на хондроцитите, причинявайки намаляване на синтеза на протеогликани и ДНК и разграждане на хрущялната матрица. Тези ефекти обикновено са съпроводени с производството на свободни кислородни радикали.
Влиянието на йонната концентрация и осмотичното налягане на околната среда върху функцията на хондроцитите
В естествения хрущял концентрацията на йони се различава значително от тази в други тъкани: съдържанието на натрий в извънклетъчната среда е 250-350 mmol, а осмоларността му е 350-450 mosmol. Когато хондроцитите се изолират от извънклетъчната мембрана (ECM) и се инкубират в стандартна среда (DMEM (Dulbecco's Minimal Essential Medium), осмоларността е 250-280,7 mosmol), средата около клетките се променя драстично. Освен това концентрацията на калций и калий в стандартната среда е значително по-ниска, отколкото в естествената тъкан, а концентрацията на аниони е значително по-висока.
Добавянето на захароза към средата повишава нейната осмоларност и индуцира преходно вътреклетъчно увеличение на концентрацията на H + и калциеви аниони в цитозола. Такива вътреклетъчни промени могат да повлияят на процесите на диференциация на хондроцитите и тяхната метаболитна активност. J. Urban et al. (1993) установяват, че включването на 35 8-сулфат и 3H -пролин от изолирани хондроцити, инкубирани в стандартна DMEM среда за 2-4 часа, е само 10% от това в нативната тъкан. Интензивността на синтеза достига максимум при осмоларност на извънклетъчната среда от 350-400 mosmol както в прясно изолирани хондроцити, така и в експлантати от хрущялна тъкан. Освен това, обемът на хондроцитите се увеличава с 30-40% след поставяне на изолираните клетки в стандартна DMEM среда с определена осмоларност. Въпреки това, при култивиране на хондроцити при условия на нефизиологична осмоларност в продължение на 12-16 часа, клетките се адаптират към новите условия, намалявайки интензивността на биосинтеза пропорционално на изместването на осмоларността на извънклетъчната среда.
П. Боргети и др. (1995) изследват ефекта на осмоларността на извънклетъчната среда върху растежа, морфологията и биосинтеза на свински хондроцити. Авторите демонстрират сходни биохимични и морфологични характеристики на хондроцитите, култивирани в среди с осмоларност 0,28 и 0,38 мосмол. При осмоларност на средата 0,48 мосмол се наблюдава намаляване на клетъчната пролиферация и протеиновия синтез през първите 4-6 часа на култивиране, но впоследствие тези параметри се възстановяват и в крайна сметка достигат контролни стойности. Когато хондроцитите се култивират в среда с осмоларност 0,58 мосмол, клетките губят способността си да поддържат физиологичната интензивност на пролиферативните процеси и след 6 дни броят на хондроцитите намалява значително. При осмоларност на средата 0,58 мосмол се наблюдава силно инхибиране на протеиновия синтез. Освен това, когато се култивират в среда с осмоларност 0,28-0,38 mOsm, хондроцитите запазват своя физиологичен фенотип; при по-висока осмоларност (0,48-0,58 mOsm) настъпват значителни промени в клетъчната морфология, което се проявява със загуба на характерния фенотип, трансформация на хондроцитите във фибробластоподобни клетки и загуба на способността на клетките да сглобяват матрични протеогликани. Резултатите от това проучване показват способността на хондроцитите да реагират на ограничени колебания в осмоларността на извънклетъчната среда.
Промените в концентрацията на други йони също могат да повлияят на процесите на биосинтез в хондроцитите. По този начин, степента на включване на 35S (сулфат) се увеличава наполовина с увеличаване на концентрацията на калиеви йони от 5 mmol (концентрацията в стандартната DM EM среда) до 10 mmol (концентрацията в ECM in vivo). Концентрации на калций под 0,5 mmol насърчават производството на колаген от зрели говежди хондроцити, докато концентрация от 1-2 mmol (съответстваща на концентрацията в стандартната DM EM среда) причинява значително намаляване на синтеза на колаген. Умерено увеличение на биосинтеза се наблюдава при високи нива на калций (2-10 mmol). Различни катиони участват в прикрепването на хондроцитите към ECM протеините. По този начин, магнезиевите и мангановите йони осигуряват прикрепване към фибронектин и колаген тип II, докато калциевите йони не участват в прикрепването на хондроцитите към протеините. По този начин, резултатите от описаните изследвания показват влиянието на промените в извънклетъчните йони на калий, натрий, калций и осмоларността на средата върху биосинтетичната функция на хондроцитите, инкубирани в стандартна среда.
Влияние на механичното натоварване върху метаболизма на хондроцитите
Обездвижването на ставите причинява обратима атрофия на хрущяла, което показва необходимостта от механични стимули за нормални метаболитни процеси в извънклетъчната мембрана (ECM). В повечето случаи използваните модели на клетъчни култури съществуват при нормално атмосферно налягане. M. Wright et al. (1996) показват, че механичната среда влияе върху метаболизма на хондроцитите, като клетъчният отговор зависи от интензивността и честотата на компресивното натоварване. Експерименти с натоварване върху експлантати от непокътнат ставен хрущял in vitro демонстрират намаляване на синтеза на протеини и протеогликани под действието на статично натоварване, докато динамичното натоварване стимулира тези процеси. Точните механизми на ефекта на механичното натоварване върху хрущяла са сложни и вероятно са свързани с клетъчна деформация, хидростатично налягане, осмотично налягане, електрически потенциал и повърхностни клетъчни рецептори за матрични молекули. За да се изследва ефектът на всеки от тези параметри, е необходимо да се създаде система, в която един параметър може да се променя независимо. Например, експлантната култура не е подходяща за изучаване на клетъчна деформация, но може да се използва за изучаване на общия ефект на налягането върху метаболитната активност на хондроцитите. Компресията на хрущяла води до клетъчна деформация и е съпроводена от появата на хидростатичен градиент на налягането, електрически потенциал, флуиден поток и промени във физикохимични параметри като съдържанието на вода в матрицата, плътността на електрическия заряд и нивото на осмотично налягане. Клетъчната деформация може да бъде изследвана с помощта на изолирани хондроцити, потопени в агарозен или колагенов гел.
Разработени са няколко системи за изследване на ефекта от механичната стимулация върху хондроцитна култура. Някои изследователи използват системи, при които налягането се прилага върху клетъчната култура през газовата фаза. Така, JP Veldhuijzen et al. (1979), използвайки налягане 13 kPa над атмосферното с ниска честота (0,3 Hz) в продължение на 15 минути, наблюдават увеличение на синтеза на cAMP и протеогликани и намаляване на синтеза на ДНК. R. Smith et al. (1996) показват, че периодичното излагане на култура от първични говежди хондроцити на хидростатично налягане (10 MPa) с честота 1 Hz в продължение на 4 часа води до увеличение на синтеза на агрекан и колаген тип II, докато постоянното налягане не влияе на тези процеси. Използвайки подобна система, Wright et al. (1996) съобщават, че цикличното налягане върху клетъчната култура е свързано с хиперполяризация на клетъчната мембрана на хондроцитите и активиране на Ca2 + -зависими калиеви канали. По този начин, ефектите на цикличното налягане се медиират от активирани от разтягане йонни канали в хондроцитната мембрана. Реакцията на хондроцитите на хидростатично налягане зависи от условията на клетъчната култура и честотата на приложеното натоварване. По този начин, цикличното хидростатично налягане (5 MPa) намалява включването на сулфат в хондроцитния монослой при честота 0,05, 0,25 и 0,5 Hz, докато при честота, по-голяма от 0,5 Hz, включването на сулфат в хрущялния експлант се увеличава.
М. Бушман и др. (1992) съобщават, че хондроцитите в агарозни гелове променят биосинтеза в отговор на статично и динамично механично натоварване по същия начин, както култивирания непокътнат орган. Авторите установяват, че механичното натоварване генерира хиперосмотичен стимул с последващо понижаване на pH в хондроцитите.
Ефектът от механичното разтягане може да бъде изследван върху клетъчна култура, потопена в гел. Силата на разтягане може да бъде създадена с помощта на компютърно контролиран вакуум. Когато системата е под определена степен на вакуум, дъното на петриевата паничка с клетъчната култура се удължава с известна величина, като деформацията е максимална по краищата на дъното на паничката и минимална в центъра. Разтягането се предава и на хондроцитите, култивирани в петриевата паничка. Използвайки този метод, К. Холмвал и др. (1995) показаха, че в хондросаркомни клетки, култивирани в колагенов (тип II) гел, експресията на mRNA на 2β-интегрин е повишена . 2β-интегринът е способен да се свързва с колаген тип II. Той се счита за механорецептор, тъй като взаимодейства с актин-свързващи протеини, като по този начин свързва извънклетъчната мембрана (ECM) и цитоскелета.
Влиянието на pH върху метаболизма на хондроцитите
PH на интерстициалната течност на извънклетъчната мембрана на хрущялната тъкан е по-киселинно, отколкото в други тъкани. А. Марудас (1980) определя pH на ставната хрущялна матрица на 6,9. Б. Диамант и др. (1966) установяват pH от 5,5 при патологични условия. Известно е, че хондроцитите живеят при ниско PO2, което показва важната роля на гликолизата (95% от целия глюкозен метаболизъм) в метаболизма на тези клетки; гликолизата е съпроводена с производството на голямо количество млечна киселина.
В допълнение към подкиселяването на средата от продуктите на гликолизата, самите компоненти на матрицата са от голямо значение. Голямото количество фиксиран отрицателен заряд върху протеогликаните променя извънклетъчния йонен състав: наблюдава се висока концентрация на свободни катиони (напр. H +, Na +, K + ) и ниска концентрация на аниони (напр. O2, HCO3). Освен това, под влияние на механично натоварване, водата се изхвърля от извънклетъчната мембрана (ECM), което води до увеличаване на концентрацията на фиксирани отрицателни заряди и привличане на повече катиони в матрицата. Това е съпроводено с намаляване на pH на извънклетъчната среда, което влияе на вътреклетъчното pH, като по този начин променя метаболизма на хондроцитите. R. Wilkin и A. Hall (1995) изследват ефекта на pH на извънклетъчната и вътреклетъчната среда върху биосинтеза на матрицата от изолирани говежди хондроцити. Те наблюдават двойна модификация на матричния синтез с намаляване на pH. Леко понижение на pH (7,4
[ 34 ], [ 35 ], [ 36 ], [ 37 ], [ 38 ], [ 39 ], [ 40 ]
Влиянието на състава на хранителната среда върху метаболизма на хондроцитите
Средата за култивиране на хондроцити трябва да съответства на експерименталните условия. През последните години за оптимизиране на условията на култивиране се използва телешки серум. При използване на серум обаче трябва да се вземат предвид редица важни моменти:
- растеж на клетки от периферията на тъканта в органни култури,
- променливост в състава на серумите от различни серии,
- наличието на неизвестни компоненти в тях,
- повишен риск от смущения и артефакти при изучаване на влиянието на различни биологични фактори върху метаболитната активност на клетките.
Пример за последното е изследване на ефекта на EGF върху хрущялните хондроцити при плъхове. EGF стимулира включването на 3H -тимидин и увеличаване на съдържанието на ДНК в културата. Този ефект е по-изразен при ниски серумни концентрации (<1%), но при високи концентрации (>7,5%) ефектът изчезва.
Добре известно е, че нивата на синтез и разграждане в DMEM, допълнена с телешки серум, са значително повишени в сравнение с in vivo условията. Разликите между in vivo и in vitro метаболизма може да се дължат на разликите между синовиалната течност и средата, в която се култивират клетките. Lee et al. (1997) култивират млади говежди хондроцити в агароза, използвайки хранителна среда, съдържаща DMEM, допълнена с 20% телешки серум и голямо количество нормална алогенна синовиална течност. Наличието на синовиална течност в средата индуцира увеличение на количеството протеогликани, до 80% от общото количество синовиална течност. Тези резултати показват, че синовиалната течност в културата индуцира ниво на метаболизъм, подобно на това in vivo, с високо ниво на синтез на гликозаминогликани и ниско ниво на клетъчно делене.
G. Verbruggen et al. (1995) показват, че синтезът на 35S -arrpeKaHa от човешки хондроцити, култивирани в агароза в безсерумна DMEM, е 20-30% от нивото на синтез, наблюдавано в DMEM, допълнена с 10% телешки серум. Авторите определят степента, до която IGF-1, IGF-2, TGF-R или инсулин възстановяват производството на агрекан в безсерумна среда. Авторите заключават, че 100 ng/ml инсулин, IGF-1 или IGF-2 частично възстановяват синтеза на агрекан до 39-53% от контролното ниво. Не се наблюдава синергизъм или кумулация при комбинация от изброените фактори. В същото време, 10 ng/ml TGF-R в присъствието на 100 ng/ml инсулин стимулира синтеза на агрекан до 90% или повече от референтното ниво. Накрая, човешкият серумен трансферин, самостоятелно или в комбинация с инсулин, не повлиява синтеза на агрекан. Когато телешкият серум беше заменен с говежди серумен албумин, съдържанието на агреканови агрегати намаля значително. Обогатяването на хранителната среда с инсулин, IGF или TGF-R частично възстанови способността на клетките да произвеждат агреканови агрегати. Освен това, IGF-1 и инсулинът са способни да поддържат хомеостазата в клетъчните култури. След 40 дни култивиране в среда, обогатена с 10-20 ng/ml IGF-1, синтезът на протеогликани се поддържаше на същото ниво или дори по-високо в сравнение със среда, съдържаща 20% телешки серум. Катаболните процеси протичаха по-бавно в среда, обогатена с IGF-1, отколкото в среда, обогатена с 0,1% разтвор на албумин, но малко по-бързо в среда, обогатена с 20% серум. В дългоживеещи култури, 20 ng/ml IGF-1 поддържа стабилно състояние на клетките.
Д. Лий и др. (1993) сравняват ефекта на състава на културалната среда (DMEM, DMEM+20% телешки серум, DMEM+20 ng/ml IGF-1) върху синтеза на ДНК в култура от експлант на хрущялна тъкан, монослойна култура и в агарозна суспензия. При култивиране в агароза в присъствието на серум, авторите наблюдават тенденция хондроцитите да се групират в големи клъстери. Клетките, култивирани без серум или с IGF-1, запазват кръгла форма в агароза, събират се на малки групи, но не образуват големи агрегати. В монослой, синтезът на ДНК е значително по-висок в среда, съдържаща серум, отколкото в среда, обогатена с IGF-1; синтезът на ДНК в последната е значително по-висок, отколкото в необогатената среда. Не са открити разлики в синтеза на ДНК, когато хондроцитите са култивирани в агарозна суспензия в необогатена среда и в среда с IGF-1. В същото време, култивирането на хондроцитни суспензии в агароза в обогатена със серум среда е съпроводено с повишено включване на радионуклеотида 3H -тимидин в сравнение с други среди.
Витамин С е необходим за активирането на ензими, участващи във формирането на стабилна спирална структура на колагеновите фибрили. Хондроцитите с дефицит на аскорбинова киселина синтезират недохидроксилирани неспирални колагенови прекурсори, които се секретират бавно. Прилагането на аскорбинова киселина (50 μg/ml) причинява хидроксилиране на колаген тип II и IX и тяхната секреция в нормални количества. Добавянето на витамин С не повлиява нивото на синтез на протеогликани. Следователно, секрецията на колаген се регулира независимо от секрецията на протеогликани.