Медицински експерт на статията

Нови публикации
Яйчник
Последно прегледани: 04.07.2025

Цялото съдържание на iLive е медицински прегледано или е проверено, за да се гарантира възможно най-голяма точност.
Имаме строги насоки за снабдяване и само свързваме реномирани медийни сайтове, академични изследователски институции и, когато е възможно, медицински проучвания, които се разглеждат от специалисти. Имайте предвид, че номерата в скоби ([1], [2] и т.н.) са линкове към тези проучвания.
Ако смятате, че някое от съдържанието ни е неточно, остаряло или под съмнение, моля, изберете го и натиснете Ctrl + Enter.
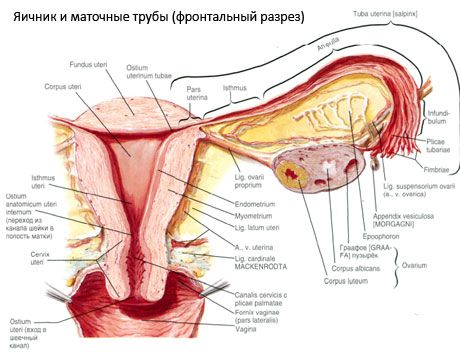
Яйчникът (ovarium; гръцки oophoron) е чифтен орган, женска полова жлеза, разположена в тазовата кухина зад широката връзка на матката. В яйчниците се развиват и узряват женските репродуктивни клетки (яйца) и се образуват женски полови хормони, които попадат в кръвта и лимфата. Яйчникът има яйцевидна форма, донякъде сплескан в предно-задната посока. Цветът на яйчника е розов. По повърхността на яйчника на родила жена се виждат вдлъбнатини и белези - следи от овулация и трансформация на жълтото тяло. Яйчникът тежи 5-8 г. Размерите на яйчника са: дължина 2,5-5,5 см, ширина 1,5-3,0 см, дебелина - до 2 см. Яйчникът има две свободни повърхности: медиалната повърхност (facies medialis), обърната към тазовата кухина, частично покрита от фалопиевата тръба, и страничната повърхност (facies lateralis), съседна на страничната стена на таза, до леко изразена вдлъбнатина - яйчниковата ямка. Тази ямка е разположена в ъгъла между външните илиачни съдове, покрити с перитонеум отгоре, и маточните и обтураторните артерии отдолу. Зад яйчника ретроперитонеално отгоре надолу преминава уретерът на съответната страна.
Повърхностите на яйчника преминават в изпъкнал свободен (задeн) ръб (margo liber), отпред - в мезентериален ръб (margo mesovaricus), прикрепен посредством къса гънка на перитонеума (мезентериума на яйчника) към задния лист на широката връзка на матката. На този преден ръб на органа има жлебова вдлъбнатина - хилумът на яйчника (hilum ovarii), през който артерията и нервите влизат в яйчника, а вените и лимфните съдове излизат. Яйчникът също има два края: заобления горен тръбен край (extremitas tubaria), обърнат към фалопиевата тръба, и долния маточен край (extremitas utenna), свързан с матката чрез яйчниковата връзка (lig. ovarii proprium). Тази връзка, във формата на кръгло връвче с дебелина около 6 мм, отива от маточния край на яйчника до страничния ъгъл на матката, разположен между двата листа на широката връзка на матката. Лигаментният апарат на яйчника включва също лигаментния суспензорий на яйчника (lig.suspensorium ovarii), който представлява гънка на перитонеума, простираща се от стената на малкия таз до яйчника и съдържаща яйчниковите съдове и снопчета от влакнести влакна вътре. Яйчникът е фиксиран от къс мезентериум (mesovarium), който представлява дупликация на перитонеума, простираща се от задния листък на широкия лигамент на матката до мезентериалния ръб на яйчника. Самите яйчници не са покрити от перитонеума. Най-големият яйчников ръб на фалопиевата тръба е прикрепен към тръбния край на яйчника. Топографията на яйчника зависи от положението на матката, нейния размер (по време на бременност). Яйчниците са много подвижни органи на малката тазова кухина.
 [
[ Съдове и нерви на яйчника
Кръвоснабдяването на яйчниците се осигурява от aa. et vv. ovaricae et uterinae. И двете яйчникови артерии (aa. ovaricae dextra et sinistra) произхождат от предната повърхност на аортата, точно под бъбречните артерии; дясната обикновено произхожда от аортата, а лявата - от бъбречната артерия. Насочвайки се надолу и странично по предната повърхност на големия псоас мускул, всяка яйчникова артерия пресича отпред уретера (отдавайки разклонения към него), външните илиачни съдове, граничната линия и навлиза в тазовата кухина, като се намира тук в суспензорния лигамент на яйчника. Следвайки в медиална посока, яйчниковата артерия преминава между листовете на широкия лигамент на матката под фалопиевата тръба, отдавайки разклонения към нея, и след това в мезентериума на яйчника; тя навлиза в хилуса на яйчника.
Клоновете на яйчниковата артерия широко анастомозират с яйчниковите клонове на маточната артерия. Венозният отток от яйчниците се осъществява предимно в яйчниковия венозен плексус, разположен в областта на яйчниковия хилум. Оттук оттокът на кръвта протича в две посоки: през маточните и яйчниковите вени. Дясната яйчникова вена има клапи и се влива в долната куха вена. Лявата яйчникова вена се влива в лявата бъбречна вена, въпреки че няма клапи.
Лимфният дренаж от яйчниците се осъществява през лимфните съдове, особено обилно в областта на органната порта, където се разграничава субовариалният лимфен плексус. След това лимфата се изхвърля по яйчниковите лимфни съдове към парааорталните лимфни възли.
Инервация на яйчниците
Симпатикова - осигурява се от постганглионни влакна от чоячния (слънчев), горния мезентериален и хипогастралния плексус; парасимпатикова - от висцералните сакрални нерви.
Структура на яйчника
Повърхността на яйчника е покрита с еднослоен зародишен епител. Под него се намира плътна съединителнотъканна протеинова обвивка (tunica albuginea). Съединителната тъкан на яйчника образува неговата строма (stroma ovarii), богата на еластични влакна. Веществото на яйчника, неговият паренхим, е разделено на външен и вътрешен слой. Вътрешният слой, разположен в центъра на яйчника, по-близо до неговата порта, се нарича медула (medulla ovarii). В този слой, в рехкава съединителна тъкан, се намират множество кръвоносни и лимфни съдове и нерви. Външният слой на яйчника - кората (cortex ovarii) е по-плътен. Съдържа много съединителна тъкан, в която са разположени зреещите първични яйчникови фоликули (folliculi ovarici primarii), вторични (везикуларни) фоликули (folliculi ovarici secundarii, s.vesiculosi), както и зрели фоликули, Граафови фоликули (folliculi ovarici maturis), както и жълти и атретични телца.
Всеки фоликул съдържа женска репродуктивна яйцеклетка или ооцит (овоцитус). Яйцеклетката е с диаметър до 150 µm, кръгла, съдържа ядро, голямо количество цитоплазма, която освен клетъчни органели съдържа протеиново-липидни включвания (жълтък), гликоген, необходими за храненето на яйцеклетката. Яйцеклетката обикновено изразходва запасите си от хранителни вещества в рамките на 12-24 часа след овулацията. Ако не настъпи оплождане, яйцеклетката умира.
Човешката яйцеклетка има две мембрани, които я покриват. Вътре се намира цитолемата, която е цитоплазмената мембрана на яйцеклетката. Отвън на цитолемата се намира слой от така наречените фоликуларни клетки, които защитават яйцеклетката и изпълняват хормонообразуваща функция - те секретират естрогени.
Физиологичното положение на матката, тръбите и яйчниците се осигурява от окачващия, фиксиращия и поддържащия апарат, който обединява перитонеума, връзките и тазовата тъкан. Окачващият апарат е представен от сдвоени образувания, той включва кръглите и широките връзки на матката, собствените връзки и окачващите връзки на яйчниците. Широките връзки на матката, собствените и окачващите връзки на яйчниците държат матката в средно положение. Кръглите връзки издърпват фундуса на матката напред и осигуряват неговия физиологичен наклон.
Фиксиращият (закотвящ) апарат осигурява позицията на матката в центъра на малкия таз и прави практически невъзможно нейното изместване встрани, напред и назад. Но тъй като лигаментният апарат се отделя от матката в долната си част, са възможни наклони на матката в различни посоки. Фиксиращият апарат включва връзки, разположени в рехавата тъкан на таза и простиращи се от долната част на матката до страничните, предната и задната стени на таза: сакромагични, кардинални, маточно-везикални и везикопубични връзки.
В допълнение към мезовариума се разграничават следните яйчникови връзки:
- суспензорният лигамент на яйчника, по-рано обозначен като инфундибулопелвичен лигамент. Той представлява гънка на перитонеума с преминаващи през нея кръвоносни съдове (a. et v. ovarica) и лимфни съдове и нерви на яйчника, опъната между страничната стена на таза, лумбалната фасция (в областта на разделянето на общата илиачна артерия на външна и вътрешна) и горния (тръбен) край на яйчника;
- Собственият лигамент на яйчника преминава между листчетата на широкия маточен лигамент, по-близо до задния лист, и свързва долния край на яйчника със страничния ръб на матката. Собственият лигамент на яйчника е прикрепен към матката между началото на фалопиевата тръба и кръглия лигамент, зад и над последния. В дебелината на лигамента преминават rr. ovarii, които са крайните клонове на маточната артерия;
- Апендикуларно-яйчникова връзка Връзката се простира от мезентериума на апендикса до десния яйчник или широката връзка на матката под формата на гънка на перитонеума. Връзката е непостоянна и се наблюдава при 1/2 - 1/3 от жените.
Поддържащият апарат е представен от мускулите и фасцията на тазовото дъно, разделени на долен, среден и горен (вътрешен) слой.
Най-мощният е горният (вътрешен) мускулен слой, представен от сдвоения мускул, който повдига ануса. Той се състои от мускулни снопчета, които се разпростират от опашната кост до тазовите кости в три посоки (пубокоцигеус, илиокоцигеус и седалищно-коцигеус). Този мускулен слой се нарича още тазова диафрагма.
Средният слой мускули е разположен между симфизата, срамната и седалищната кости. Средният слой мускули - урогениталната диафрагма - заема предната половина на тазовия изход, през който преминават уретрата и влагалището. В предния отдел между нейните листове се намират мускулни снопчета, които образуват външния сфинктер на уретрата, а в задния отдел - мускулни снопчета, които се простират в напречна посока - дълбокият напречен мускул на перинеума.
Долният (външен) слой на мускулите на тазовото дъно се състои от повърхностни мускули, чиято форма наподобява числото 8. Те включват булбокавернозен, ишиокавернозен, външен анален сфинктер и повърхностен напречен перинеален мускул.
Онтогенеза на яйчниците
Процесът на растеж и атрезия на фоликулите започва на 20-та седмица от бременността и до момента на раждането в яйчниците на момичето остават до 2 милиона яйцеклетки. До менархе броят им намалява до 300 хиляди. През целия период на репродуктивен живот не повече от 500 фоликула достигат зрялост и овулират. Първоначалният растеж на фоликулите не зависи от стимулация с FSH, е ограничен и атрезията настъпва бързо. Смята се, че вместо стероидни хормони, локалните автокринни/паракринни пептиди са основният регулатор на растежа и атрезията на първичните фоликули. Смята се, че процесът на растеж и атрезия на фоликулите не се прекъсва от никакви физиологични процеси. Този процес продължава във всички възрасти, включително вътрематочния период и менопаузата, и се прекъсва от бременност, овулация и ановулация. Механизмът, който задейства растежа на фоликулите и техният брой във всеки специфичен цикъл, все още не е ясен.
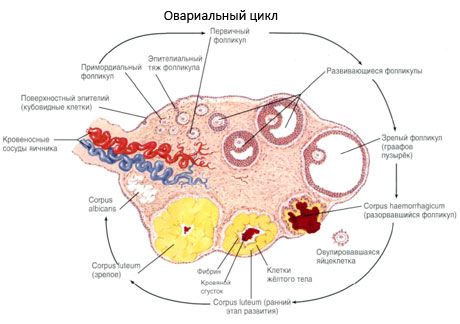
По време на своето развитие фоликулът преминава през няколко етапа на развитие. Първичните зародишни клетки произхождат от ендодермата на жълтъчния сак, алантоиса, и мигрират към гениталната област на ембриона на 5-6-та седмица от бременността. В резултат на бързото митотично делене, което продължава от 6-8-ма до 16-20-та седмица от бременността, в яйчниците на ембриона се образуват до 6-7 милиона ооцити, заобиколени от тънък слой гранулозни клетки.
Преантрален фоликул - ооцитът е обграден от мембрана (Zona pellucida). Гранулозните клетки, обграждащи ооцита, започват да пролиферират, като растежът им зависи от гонадотропините и корелира с нивото на естрогените. Гранулозните клетки са мишена за FSH. На етапа на преантралния фоликул гранулозните клетки са способни да синтезират три класа стероиди: предимно индуцира активността на ароматазата, основният ензим, който превръща андрогените в естрадиол. Смята се, че естрадиолът е способен да увеличи броя на собствените си рецептори, упражнявайки директен митогенен ефект върху гранулозните клетки, независимо от FSH. Счита се за паракринен фактор, който усилва ефектите на FSH, включително активирането на процесите на ароматизация.
FSH рецепторите се появяват върху мембраните на гранулозните клетки веднага щом започне растежът на фоликула. Намаляването или увеличаването на FSH води до промяна в броя на неговите рецептори. Това действие на FSH се модулира от растежни фактори. FSH действа чрез G-протеина, аденилат циклазната система, въпреки че стероидогенезата във фоликула се регулира главно от FSH, в този процес участват много фактори: йонни канали, тирозин киназни рецептори, фосфолипазна система на вторични посланици.
Ролята на андрогените в ранното развитие на фоликулите е сложна. Гранулозните клетки имат андрогенни рецептори. Те са не само субстрат за индуцирана от FSH ароматизация до естрогени, но могат също така да усилят ароматизацията при ниски концентрации. Когато нивата на андрогени се повишат, преантралните гранулозни клетки предпочитат да не избират пътя на ароматизация до естрогени, а по-простия андрогенен път чрез 5α-редуктаза, за да се превърнат в андроген, който не може да се превърне в естроген, като по този начин инхибират ароматазната активност. Този процес също така инхибира образуването на FSH и LH рецептори, като по този начин спира развитието на фоликулите.
Процесът на ароматизация, фоликулът с високи нива на андрогени претърпява процеси на атрезия. Растежът и развитието на фоликула зависи от способността му да превръща андрогените в естрогени.
В присъствието на FSH, доминиращото вещество на фоликуларната течност ще бъдат естрогените. При липса на FSH - андрогените. LH обикновено липсва във фоликуларната течност до средата на цикъла. Веднага щом нивото на LH се повиши, митотичната активност на гранулозните клетки намалява, появяват се дегенеративни промени и нивото на андрогени във фоликула се повишава. Нивото на стероиди във фоликуларната течност е по-високо, отколкото в плазмата, и отразява функционалната активност на яйчниковите клетки: гранулозни и тека клетки. Ако единствената цел за FSH са гранулозните клетки, тогава LH има много цели - тека клетки, стромални и лутеинови клетки и гранулозни клетки. Както гранулозните, така и тека клетките имат способността за стероидогенеза, но ароматазната активност преобладава в гранулозните клетки.
В отговор на LH, тека клетките произвеждат андрогени, които след това се превръщат от гранулозните клетки в естрогени чрез FSH-индуцирана ароматизация.
С растежа на фоликула, тека клетките започват да експресират гени за LH рецептори, P450 sec и 3β-хидроксистероид дехидрогеназа, инсулиноподобният растежен фактор (IGF-1) синергично с LH увеличава генната експресия, но не стимулира стероидогенезата.
Овариалната стероидогенеза винаги е LH-зависима. С растежа на фоликула, тека клетките експресират ензима P450c17, който образува андроген от холестерол. Гранулозните клетки нямат този ензим и са зависими от тека клетките, за да произвеждат естрогени от андрогени. За разлика от стероидогенезата, фоликулогенезата е FSH-зависима. С растежа на фоликула и повишаването на нивата на естроген се активира механизъм за обратна връзка - производството на FSH се инхибира, което от своя страна води до намаляване на ароматазната активност на фоликула и в крайна сметка до фоликуларна атрезия чрез апоптоза (програмирана клетъчна смърт).
Механизмът на обратна връзка на естрогените и FSH инхибира развитието на фоликулите, които са започнали да растат, но не и на доминантния фоликул. Доминантният фоликул съдържа повече FSH рецептори, които подпомагат пролиферацията на гранулозни клетки и ароматизацията на андрогените до естрогени. Освен това, паракринните и автокринните пътища действат като важен координатор на развитието на антралните фоликули.
Автокринният/паракринният регулатор се състои от пептиди (инхибин, активин, фолистатин), които се синтезират от гранулозни клетки в отговор на FSH и навлизат във фоликуларната течност. Инхибинът намалява секрецията на FSH; активинът стимулира освобождаването на FSH от хипофизната жлеза и усилва действието на FSH в яйчника; фолистатинът потиска активността на FSH, вероятно чрез свързване с активин. След овулацията и развитието на жълтото тяло, инхибинът е под контрола на LH.
Растежът и диференциацията на яйчниковите клетки се влияят от инсулиноподобни растежни фактори (IGE). IGF-1 действа върху гранулозните клетки, причинявайки повишаване на цикличния аденозин монофосфат (цАМФ), прогестерона, окситоцина, протеогликана и инхибина.
IGF-1 действа върху тека клетките, причинявайки повишено производство на андрогени. Тека клетките, от своя страна, произвеждат тумор некрозис фактор (TNF) и епидермален растежен фактор (EGF), които също се регулират от FSH.
EGF стимулира пролиферацията на гранулозни клетки. IGF-2 е основният растежен фактор във фоликуларната течност, а в нея се откриват още IGF-1, TNF-a, TNF-3 и EGF.
Нарушаването на паракринната и/или автокринната регулация на яйчниковата функция изглежда играе роля в нарушенията на овулацията и в развитието на поликистозни яйчници.
С растежа на антралния фоликул, съдържанието на естроген във фоликуларната течност се увеличава. В пика на тяхното увеличение, рецепторите за LH се появяват върху гранулозните клетки, настъпва лутеинизация на гранулозните клетки и се увеличава производството на прогестерон. По този начин, в преовулаторния период, увеличаването на производството на естроген причинява появата на LH рецептори, LH, от своя страна, причинява лутеинизация на гранулозните клетки и производство на прогестерон. Увеличаването на прогестерона намалява нивата на естроген, което очевидно причинява втори пик на FSH в средата на цикъла.
Смята се, че овулацията настъпва 10-12 часа след пика на LH и 24-36 часа след пика на естрадиола. Смята се, че LH стимулира редукционното делене на ооцита, лутеинизацията на гранулозните клетки и синтеза на прогестерон и простагландин във фоликула.
Прогестеронът усилва активността на протеолитичните ензими, които заедно с простагландин участват в разкъсването на фоликулната стена. Индуцираният от прогестерон пик на FSH позволява на ооцита да излезе от фоликула чрез превръщане на плазминогена в протеолитичния ензим плазмин и осигурява достатъчен брой LH рецептори за нормалното развитие на лутеалната фаза.
В рамките на 3 дни след овулацията гранулозните клетки се увеличават по размер и в тях се появяват характерни вакуоли, пълни с пигмент лутеин. Тека-лутеалните клетки се диференцират от теката и стромата и стават част от жълтото тяло. Капилярите, проникващи в жълтото тяло, се развиват много бързо под влияние на фактори на ангиогенезата и с подобрена васкуларизация се увеличава производството на прогестерон и естрогени. Активността на стероидогенезата и продължителността на живота на жълтото тяло се определят от нивото на LH. Жълтото тяло не е хомогенно клетъчно образувание. Освен 2 вида лутеални клетки, то съдържа ендотелни клетки, макрофаги, фибробласти и др. Големите лутеални клетки произвеждат пептиди (релаксин, окситоцин) и са по-активни в стероидогенезата с по-голяма ароматазна активност и по-голям синтез на прогестерон, отколкото малките клетки.
Пикът на прогестерона настъпва на 8-ия ден след пика на LH. Отбелязва се, че прогестеронът и естрадиолът се секретират епизодично в лутеалната фаза в корелация с пулсиращата продукция на LH. С образуването на жълтото тяло, контролът върху производството на инхибин преминава от FSH към LH. Инхибинът се увеличава с повишаването на естрадиола преди пика на LH и продължава да се увеличава след пика на LH, въпреки че нивата на естроген намаляват. Въпреки че инхибинът и естрадиолът се секретират от гранулозните клетки, те се регулират по различни пътища. Спадът на инхибина в края на лутеалната фаза допринася за повишаване на FSH за следващия цикъл.
Жълтото тяло намалява много бързо - на 9-11-ия ден след овулацията.
Механизмът на дегенерация е неясен и не е свързан с лутеолитичната роля на естрогените или с рецепторен механизъм, както се наблюдава в ендометриума. Има и друго обяснение за ролята на естрогените, произвеждани от жълтото тяло. Известно е, че естрогените са необходими за синтеза на прогестеронови рецептори в ендометриума. Естрогените в лутеалната фаза вероятно са необходими за свързаните с прогестерона промени в ендометриума след овулация. Неадекватното развитие на прогестероновите рецептори, в резултат на неадекватните нива на естроген, е вероятно допълнителен механизъм за безплодие и ранни спонтанни бременности, друга форма на дефицит на лутеалната фаза. Смята се, че продължителността на живота на жълтото тяло се установява по време на овулацията. И със сигурност ще регресира, ако не се поддържа от човешки хорионгонадотропин поради бременност. По този начин, регресията на жълтото тяло води до намаляване на нивата на естрадиол, прогестерон и инхибин. Намаляването на инхибина премахва неговия потискащ ефект върху FSH; Намаляването на естрадиола и прогестерона позволява много бързо възстановяване на секрецията на GnRH и премахване на механизма за обратна връзка от хипофизната жлеза. Намаляването на инхибина и естрадиола, заедно с повишаване на GnRH, води до превишаване на FSH над LH. Увеличаването на FSH води до растеж на фоликулите с последващ избор на доминантен фоликул и започва нов цикъл, ако не настъпи бременност. Стероидните хормони играят водеща роля в репродуктивната биология и общата физиология. Те определят човешкия фенотип, влияят на сърдечно-съдовата система, костния метаболизъм, кожата, общото благосъстояние и играят ключова роля при бременността. Действието на стероидните хормони отразява вътреклетъчните и генетичните механизми, които са необходими за предаване на извънклетъчен сигнал към клетъчното ядро, за да се предизвика физиологичен отговор.
Естрогените дифузно проникват през клетъчната мембрана и се свързват с рецептори, разположени в клетъчното ядро. След това рецепторно-стероидният комплекс се свързва с ДНК. В целевите клетки тези взаимодействия водят до генна експресия, протеинов синтез и специфична клетъчна и тъканна функция.